
A short evolutionary journey across the HERC Ubiquitin Ligases
Enrico Bracco2*, Cristina Panuzzo1, Barbara Pergolizzi1*
1Dept. of Clinical & Biological Sciences, University of Turin, Italy
2Dept. of Oncology, University of Turin, Italy
Abstract
HECT ubiquitin ligases are key components of the eukaryotic ubiquitin-proteasome system controlling different cellular physiological aspects as well as the genesis of several human diseases. Among the HECT family, the HERC subfamily members are characterized by having one or more RCC1-like domains, a C-terminal HECT domain and the molecular mass ranging approximately from 120 kDa to 500 kDa. Due to their large size, some of them are refractory to functional characterization. We have recently identified and functionally characterized a novel large HECT member in Dictyostelium discoideum that, in many aspects, exhibits structural similarities with the mammalian large HERC1. In the present minireview, we shortly summarize and revise the current phylogenetic history of HERC proteins among the different living organisms.
Introduction
Ubiquitination, which is the covalent ligation of ubiquitin to a substrate protein, is associated with almost every cellular process including signal transduction, DNA damage repair, cell cycle regulation and autophagic clearance. The process involves three sequential steps each of which is catalysed by a different class of enzymes. In the first step the ubiquitin is activated, in an ATP-dependent way, by the ubiquitin activating enzyme (E1). Afterwards the activated ubiquitin is physically transferred to the ubiquitin conjugating enzyme (E2). Finally, E3 ligases carry out the final step in the ubiquitination cascade, catalysing the transfer of ubiquitin from an E2 enzyme to form a covalent bond with a substrate lysine1 (Figure 1).
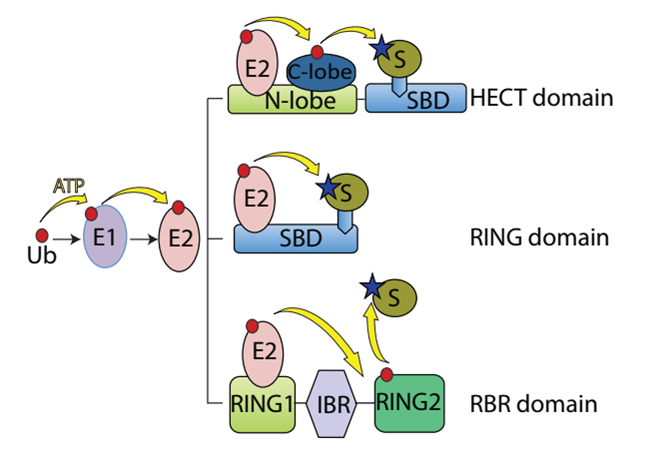
Figure 1: Schematic representation of the ubiquitin system
Ubiquitination proceeds through an initiation step during which the ubiquitin is activated by an E1 enzyme. Then, the activated ubiquitin is transferred to E2. Selectivity of substrate comes from the E3 ubiquitin ligases, which couple with E2 to bind substrates and catalyse the transfer of the activated ubiquitin from the E2 itself to the substrate. E3 HECT, RING and RBR ligase domains, act through different reaction mechanisms. While the E3-HECT ligase catalyses by itself the ubiquitin transfer to the substrate by forming an E3-ubiquitin thioester intermediate, E3-RING ligase domain does not possess direct catalytic activity but rely on the enzymatic activity of E2 to ubiquitinate the substrates. Eventually, RBR ligases combine features of both RING- and HECT-type ligases.
Three distinct families of E3 ligases have been identified, which stimulate the ubiquitin transfer either through a direct or an indirect mechanism, named RING (Really Interesting New Gene), HECT (Homologous to the E6AP Carboxyl Terminus) and RBR (Ring Between Ring)2 .
In human beings RING is the largest family with more than 600 putative members that do not have a direct catalytic role in protein ubiquitination but basically, they act as scaffold facilitating the interaction between E2 and the substrates. In contrast to RING E3 ligases, the HECT and RBR family members catalyse the ubiquitin transfer to the substrate through a two-step reaction: ubiquitin is first transferred to a catalytic cysteine residue on the E3 and then covalently linked to the substrate3,4. HECT E3s can either function on their own or in conjunction with accessory or adaptor protein5. While the selectivity and specificity for the substrate recognition for RING E3 ligase is mainly accomplished by the requirement of multisubunit complexes, for the HECT and RBR members the N-terminal region is most likely the sole that serves as the substrate recognition domain.
Despite the efforts made in the identification of potential substrate proteins of individual HECT, the physiological relevance of many of these interactions remain still unclear. Besides the animal kingdom, where the functional characterization of HECT E3 members is however available only for a handful of them, the evolutionary history of the HECT E3s and their biological roles remain still mostly unknown. In this respect we have recently characterized a novel large HECT member which might be a functional, but not structural, distant relative of the large HERC E3 sub-family6,7. In the present minireview, we will attempt to shortly summarize and revise the long evolutionary history of the HERC E3 ubiquitin ligases members.
HERC family members structural features and domain composition throughout the evolution
The structurally related HERC E3 ubiquitin ligases belong to a subfamily of the HECT E3 family. The HERC members are proteins of different size (ranging from 100-120 kDa to >500 kDa), characterized by the presence of two conserved domains: I) a catalytic HECT domain localized at their C-terminus and II) one or more (up to three) RCC1-like domain (RLD)8. HECT is a domain of ~350 amino acids firstly described in Human Papilloma Virus (HPV) E6-Associated Protein (E6AP). The HECT domain is found in different species ranging from yeast to humans. Structurally, it consists of two lobes, a larger N-terminal- (N-lobe) and a smaller C-terminal- (C-lobe) lobe connected by a short flexible linker essential for catalysis9. The C-lobe contains a crucial catalytic cysteine residue and interacts with ubiquitin during the transfer reaction while the N-lobe interacts with the E2 enzymes10. Substrates recognition is mediated by domains localized within the extended region N-terminal to the catalytic HECT domain. According to their size HERC members have been catalogued into small (HERC3-6) and large HERC (HERC1-2)11, but this classification is purely operational and not based on evolutional considerations.
The elaboration of a phylogenetic tree with the sequences of all published animal HECTs from different species suggested that the aforementioned classification was, to some extent, oversimplified12. Indeed, the phylogenetic analysis established that the small and large HERC subfamilies are only distantly related. In addition, the study allowed to trace the oldest common ancestral HERC encoding gene in the choanoflagellate Monosiga brevicollis whose genome encodes for a small HERC, mostly like the mammalian HERC4 and HERC3, but not for canonical large HERC. Animal large HERC members, namely HERC1 and HERC2, are giant proteins with molecular mass ≥500 kDa. HERC1 is structurally characterized by a HECT domain localized at its C-terminus and by a single SPRY (spl A and Ryanodine Receptor), WD40 (G protein β subunit like repeats) and two RLDs. The architecture of HERC2 is more composite than that of HERC1 because it contains multiple conserved regions other than the HECT and RLD domains, including a Cytb5 (cytochrome b5-like) region, a DOC (DOC1/APC10), a mind-bomb/HERC2 (MIB), a Cullin7 protein homology (CPH) and a ZZ-type zinc finger domain.
Recently, in a genetic suppressor screening using as parental strain a Dictyostelium discoideum TORC2 deficient mutant13 we identified, and functionally characterized, a giant HECT protein regulating some aspects of cell-communications and signal transduction. As a matter of fact, this giant E3-ubiquitin ligase controls G-protein mediated cell-to-cell signaling and acts as a suppressor for the TORC2-dependent PKB activity6. The protein displays the highest homology to the human large HERC1 (57% similarity and 38% identity within the HECT domain). Both the Dictyostelium and the human protein possess a single SPRY domain, likely mediating protein interaction, typical of several proteins and peculiar of HERC1. Furthermore, the comparison of the Dictyostelium giant E3-ubiquitin ligase with the human HERC1, shows that the first lacks the RLD whose molecular function in HERC proteins is still not fully understood. Indeed, RLD1 domain of the human HERC1 may act either as a guanine exchange factor (GEF) and guanine releasing factor (GRF) for ARF1, and as GEF for some Rab family GTPases (Rab3a, Rab5)14. On the contrary, this biochemical feature is not shared with the structurally related RLD domain of the small HERCs15. Additionally, it has been reported that the RLD1 GRF activity requires the presence of phosphatidylinositol-4,5-bisphosphate [PI(4,5)P2] indicating that the RLD1 recognizes, and non-covalently binds, phospholipids14. Remarkably, besides the HECT and SPRY domain, the Dictyostelium giant HECT harbours a Plekstrin Homology (PH) domain that represents the paradigmatic phospholipids recognition motif. To our knowledge it was the first large HECT E3-ubiquitin ligase possessing a PH domain. Thus, the protein has been named HectPH1. This unique feature, shared with the other members of the amoebozoa group (e.g. Dictyostelium sp., Acytostelium, Polysphondillium and Tieghemostelium) places the Dictyostelium HectPH1 structurally distant from canonical large HERC but potentially, functionally closer than foreseen. Currently, whether the structurally related HERC proteins, as such, have evolutionary emerged rather late, or not, remains a debated issue. Here, we attempt to shortly review the evolutionary history of HERC E3-ubiquitin ligase going back to lower eukaryotes (protists), plants and primitive algae (red and brown). As below will be briefly discussed, a database revision of the current annotated genomes highlights that the phylogenetic history of HERC proteins needs to be up-dated and slightly revised.
Indeed, thanks to the accessibility to more sequenced and annotated genomes the scenario rapidly changed and structurally related HERC encoding genes have been identified either in the protist and plant kingdoms16, as it is summarized in Table 1.
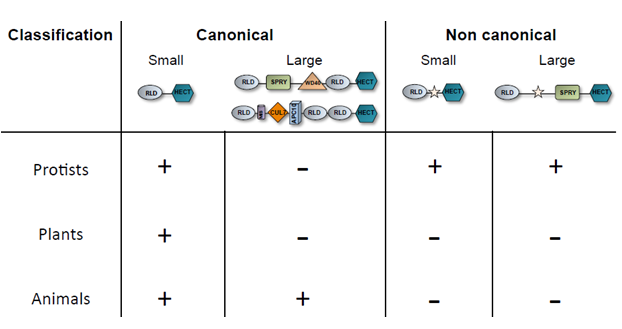
Table 1-HERC proteins distribution among the protist, plant and animal kingdoms. The yellow star in the non-canonical HERC group indicates any single, or multiple (in various order and number), domain/s in addition to the RLD, SPRY and HECT that are shared with the canonical HERCs.
Such evidence indicates that the most primitive prototype of the HERC E3-ubiquitin ligase date back to a kingdom different from that of an animal as hitherto believed12. Indeed, the vast majority of protists organisms harbour genes encoding for both putative small and large HERC. Interestingly, the structural domain composition of the N-terminal region of the vast majority of the protists putative large HERC evokes that of canonical mammalian HERC1 because they possess at least a RLD, SPRY, a single HECT and in some cases, an UBA domain but all of them lack a WD40 motif hence named “HERC-like”16. Animals such as the urochordate (Ciona intestinalis) and the cnydaria (Hydra vulgaris) harbour HERC1 proteins that, structurally, are already very close to those of mammals, albeit characterized by a single RLD instead of the canonical two archetypal of the mammalian HERC1. Noteworthy, some protists such as amoebozoa, dinoflagellates and apicomplexa encode for small HERC-like proteins, that is their size range between 120 to 200 kDa, but, besides the canonical RLD and Hect, they possess a third additional domain displaying homology either with the serine/threonine kinase, tetratricopeptide or cytochrome-C motif, respectively.
Remarkably, canonical small HERC encoding genes are found also in some red and brown algae (e.g. Fistulifera solaris, Galdieria sulphuraria and Cyanidioschyzon merolae) as well as in green plants.
The latter organisms uniquely encode for canonical small HERCs and their presence is restricted to the green algae and herbaceous plants, though the latter organisms uniquely encode for canonical small HERC members.
It appears clear that the current HERCs phylogenetic history needs to be re-assessed. Actually, we are aware that HERC proteins are not exclusively restricted to the animal kingdom but evolutionary they emerged earlier, an occurrence that is witnessed by their appearance in protist and plant kingdoms. The molecular mechanisms controlling the HERC ubiquitin ligases, as well as their enzymatic regulation and substrate specificity, are so far open questions and answering to them will represent the major future challenge to fully understand their functional roles. In this regard, the large HERCs possess a variety of motifs whose function remains currently mysterious. Besides the RLD domain, the SPRY motif is highly conserved throughout the evolution suggesting a strictly maintained role. Currently, whether the N-terminal domain of large HERC contribute to the specificity of the substrate recognition, to the enzymatic regulation or both and to a what extent to date remains unknown. The presence of HERC encoding genes in simple, and genetically amenable, organisms (e.g., amoebozoa) potentially make them very valuable and attractive models, alternative to animals, to better understand and clarify the functional roles, substrates, physical/genetic interactors and cellular substrates of these peculiar group of E3 ubiquitin ligases.
References
- Buetow L, Huang DT. Structural insights into the catalysis and regulation of E3 ubiquitin ligases. Nat Rev Mol Cell Biol 2016Oct; 17(10): 626:42.
- Metzger MB, Hristova VA, Weissman AM. HECT and RING finger families of E3 ubiquitin ligases at a glance. J Cell Sci. 2012 Feb 1; 125: 531-7.
- Wenzel DM, Klevit RE. Following Ariadne’s thread: a new perspective on RBR ubiquitin ligases. BMC Biol. 2012; 10(24).
- Huibregtse JM, Scheffner M, Beaudenon S, et al. A family of proteins structurally and functionally related to the E6-AP ubiquitin-protein ligase. Proc Natl Acad Sci USA. 1995; 92: 2563–7.
- Gatti M, Pinato S, Maiolica A, et al. RNF168 promotes noncanonical K27 ubiquitination to signal DNA damage. Cell Rep. 2015; 10(2): 226-38.
- Pergolizzi B, Bracco E, Bozzaro S. A new HECT ubiquitin ligase regulating chemotaxis and development in Dictyostelium discoideum. J Cell Sci. 2017; 1;130(3): 551:62.
- Pergolizzi B, Bozzaro S, Bracco E, and G-PDST, E2180. UiDIJMSOp, PMCID: diRPPPC, et al. G-Protein Dependent Signal Transduction and Ubiquitination in Dictyostelium. Int J Mol Sci. 2017; Oct 19: 18(0).
- Sánchez-Tena S, Cubillos-Rojas M, Schneider T, et al. Functional and pathological relevance of HERC family proteins: a decade later. Cell Mol Life Sci. 2016; 73(10): 1955-68.
- Verdecia MA, Joazeiro CA, Wells NJ, et al. Conformational flexibility underlies ubiquitin ligation mediated by the WWP1 HECT domain E3 ligase. Mol Cell. 2003; 11: 249-59.
- Huang L, Kinnucan E, Wang G, et al. Structure of an E6AP-UbcH7 complex: insights into ubiquitination by the E2-E3 enzyme cascade. Science. 1999; Nov 12(286(5443)): 1321:6.
- Hochrainer K, Mayer H, Baranyi U, et al. The human HERC family of ubiquitin ligases: novel members, genomic organization, expression profiling, and evolutionary aspects. Genomics. 2005 Feb; 85(2): 153:64.
- Marín I. Animal HECT ubiquitin ligases: evolution and functional implications. BMC Evol Biol. 2010 Feb 22;10: 56.
- Pergolizzi B, Peracino B, Silverman J, et al. Temperature-sensitive inhibition of development in Dictyostelium due to a point mutation in the piaA gene. Dev Biol. 2002; 251: 18-26.
- Garcia-Gonzalo FR, Bartrons R, Ventura F, et al. Requirement of phosphatidylinositol-4,5-bisphosphate for HERC1-mediated guanine nucleotide release from ARF proteins. FEBS Lett. 2005Jan 17; (17;579(2)): 342:8.
- Rotin D, Kumar S. Physiological functions of the HECT family of ubiquitin ligases. Nat Rev Mol Cell Biol. 2009; 10(6): 398-409.
- Grau-Bové X, Sebé-Pedrós A, Ruiz-Trillo I. A genomic survey of HECT ubiquitin ligases in eukaryotes reveals independent expansions of the HECT system in several lineages. Genome Biol Evol. 2013; 5(5): 833-47.